
花粉是植物的雄配子体,而有活力的花粉是维持植物繁衍和作物产量的基础。近日,我室室常芳团队在JIPB在线发表了题为“CLE19 Suppresses Brassinosteroid Signaling Output via the BSL-BIN2 Module to Maintain BES1 Activity and Pollen Exine Patterning in Arabidopsis ”的研究论文。该研究定义了一个全新的双信号机制,小孢子来源的 CLE19 小肽信号与绒毡层细胞的BR激素信号通过拮抗作用形成“刹车”-“油门”机制,共同保障花粉外壁(exine)的正常发育和生殖稳态。

花粉外壁是维持雄性育性和植物繁殖成功的关键结构,既能抵御外界环境胁迫,又承担着花粉-柱头识别等重要功能。其精巧外壁结构依赖于花粉母细胞/小孢子与绒毡层细胞之间的精妙交流。一旦调控失衡,就会导致花粉败育,直接威胁作物产量与繁殖能力。常芳教授团队前期发现,绒毡层通过细胞内部一系列精密调控的“前馈”-“正反馈”-“刹车”机制来维持花粉壁发育的稳态:一方面,绒毡层细胞中的门控转录因子DYT1与其下游的bHLH010/089/091转录因子间通过“前馈”和“正反馈”环路精准而迅速地启动绒毡层中孢粉素前体物质的合成(Zhu et al., 2015; Cui et al., 2016);与此同时,咖啡酰辅酶A-O-甲基转移酶1(CCoAOMT1)通过翻译后水平调控bHLH010/bHLH089的出核和降解,从而降低其活性(Lai et al., 2024)。为了防止绒毡层转录网络过度激活而导致的孢粉素前体物质过量堆积,小孢子来源的CLE19 及其同源小肽信号则作为反馈信号,通过激活其位于绒毡层细胞膜上的PXL1-SERKs受体-共受体复合物,将绒毡层转录网络的水平维持在正常水平,从而最终保障花粉正常发育(Wang et al., 2017; Yu et al., 2023, 2025)。
那么,细胞外信号CLE19 小肽及其受体共受体如何完成从膜到核的信号传递?在这一过程中CLE小肽激素与其它传统激素信号有无信号交互呢?课题组近日发表在 JIBP的研究工作,回答了上述问题:小孢子分泌的CLE19小肽及其受体PXL1通过调控一个被大家广为熟知的 BSLs-BIN2-BES1 功能模块而发挥作用。与 BR 通过 BSU1抑制BIN2进而使 BES1 维持在细胞核内的非磷酸化活性状态不同,CLE19 及其受体则通过BSU1-like 1/2/3(简称BSL1/2/3)-BIN2模块来激活转录因子BES1 的S219和S223位点的磷酸化“标签”,进而促进其出细胞核降解,并最终抑制花粉壁合成相关下游基因表达。BR 和 CLE19两种信号一个作为“油门”激活BES1 活性;另一个则作为“刹车”抑制其活性,两者的动态平衡共同保障花粉壁正常发育。
该研究首先通过定量磷酸蛋白组分析,发现 CLE19 会影响多个BR 信号转导组分的磷酸化,包括 BSL 型磷酸酶 BSL1/2/3、GSK3 样激酶 BIN2 和转录因子 BES1。为了阐释两条信号途径的相关性,课题组首先分析了 BR受体 BRI1突变体的表型,并与CLE19 过表达和功能失活型遗传材料的表型进行对比分析。有意思的是, 花粉壁观察和分子表型证据均表明BR 信号传导受阻(bri1 突变体)表现出和CLE19 过表达(CLE19-OX)一致、而和CLE19 功能失活(Dominant Negative-CLE19,DN-CLE19)相反的表型,暗示了暗示了两个信号不仅均参与花粉壁发育过程,且作用方向相反。进一步的转录组分析也支持了这个结论:在 428 个BR和 CLE19 共同的核心下游基因中,86.7% 被两信号反向调控(图 1)。
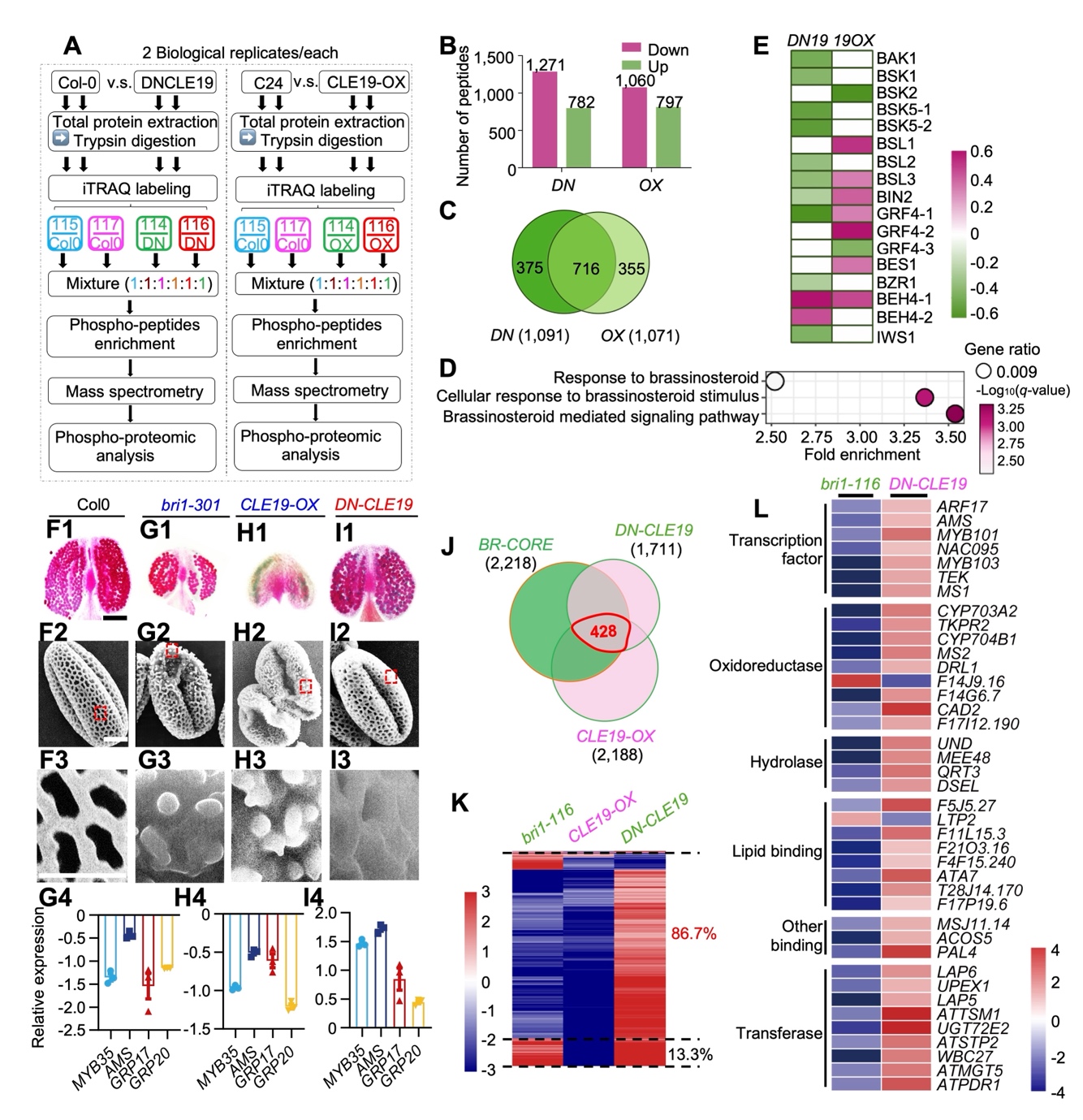
图 1. CLE19 与 BR 在花粉壁发育中拮抗发挥功能。
那么,CLE19 小肽信号和 BR 激素信号是如何拮抗调控花粉发育的呢?课题组进一步研究发现, CLE19小肽并不影响 BR 的合成,也不破坏 BR-BRI-SERK1 配基受体复合物的活性。而是通过 CLE19受体 PXL1 招募BSL1/2/3 并激活后者的磷酸化,进而通过依赖BSL1/2/3 和 BIN2 的方式激活 BES1 两个丝氨酸残基 (S219 /S223 )的磷酸化。且所有激活 BR 输出的(BR-output )BR 信号途径元件相关突变体,均表现出和 CLE19 功能缺失的表型;而所有 BR 输出被抑制的突变体则均表现出 CLE19 信号过表达的表型 (图 2)。通过构建不能磷酸化(phosphor-dead,S-A)和模拟磷酸化(phosphor-mimic,S-D) 两种形式的BES1 变体,并转基因植株进行功能分析,课题组进一步证实了依赖于 CLE19 的磷酸化激活了 BES1 的核输出和降解,抑制了花粉外植体花型所需的 BR 响应转录输出,从而最终抑制下游与花粉壁发育相关关键转录因子 MYB35、AMS 及合成酶基因的表达。
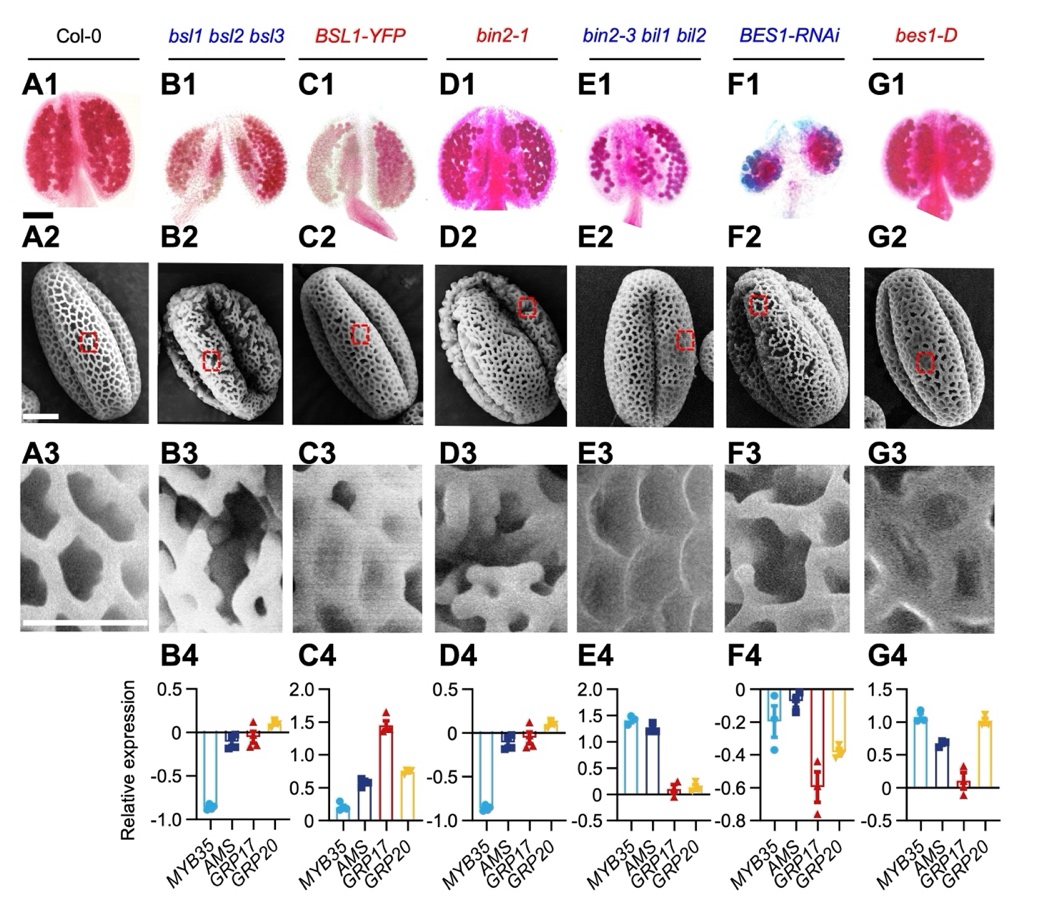
图 2. BSLs-BIN2-BES1 模块是花粉壁正常发育所必需的。
综上所述,团队的研究结果定义了一个全新的双信号机制,其中 CLE19 小肽由发育中的花粉产生,并作为一种可扩散的“分子制动剂”,通过 BSL2-BIN2-BES1信号传导轴抑制 BR 信号的输出,并最终抑制绒毡层转录网络的过度激活,确保绒毡层功能的稳定性。该工作将小孢子产生的多肽与绒毡层内的 BR 激素信号紧密联系,从机理上揭示了植物生殖过程中如何通过细胞间信号整合来维持发育的稳健性。
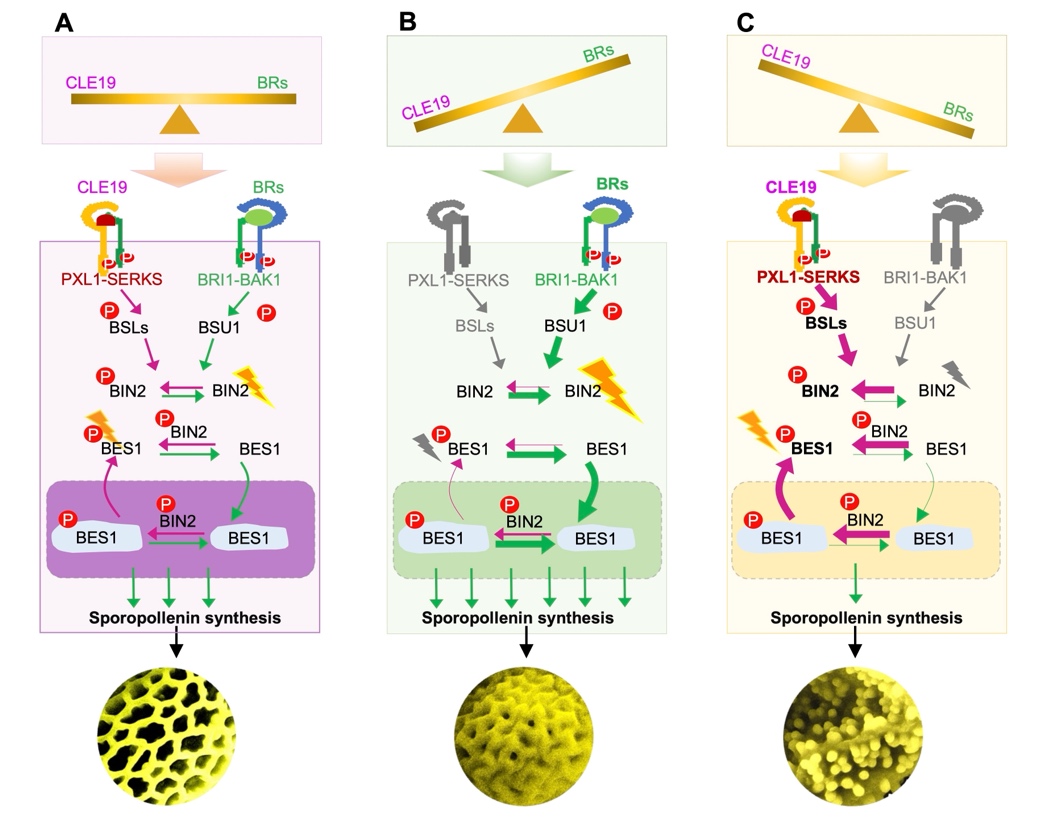
Figure 3. CLE19 与 BR 共同调控花粉发育稳态的工作模型。
复旦大学生命科学学院博士后王双双(现华东师范大学副研究员)、博士研究生张诗婷、博士后余英为论文的共同第一作者,常芳教授为通讯作者。中国农业大学宋文教授、美国罗格斯大学董娟教授、广州大学黎家教授、北京大学刘春明教授、美国宾州州立大学马红教授、复旦大学博士后叶娟英、复旦大学博士研究生王健正、刘叶乔,硕士生王静雅、李梦雨等也参与了该研究。研究工作得到了科技部重点研发计划、国家自然科学基金以及博士后科学基金的支持。
原文链接:https://doi.org/10.1111/jipb.70024

版权所有:复旦大学复杂性状的遗传调控全国重点实验室
Copyright: State Key Laboratory of Genetics and Development of Complex Phenotypes